The Invasion of Phellodendron amurense into the Urban and Suburban Woodlands of the New York City Region
by Eric C. Morgan and Jon A. Borysiewicz
Bartlett Arboretum, 151 Brookdale Rd., Stamford, CT 06903
Abstract
Phellodendron amurense Rupr. is an introduced, exotic tree species from eastern Asia which has been used in ornamental horticulture in urban and suburban areas throughout the Northeast. In recent years, even as more focus has been placed upon the impact of invasive species, Phellodendron amurense and other non-native species have continued to be planted, and little or no examination of their potential impact and spread has been undertaken. In 2008 and 2009, surveys were performed at sites where Phellodendron amurense was reported to have been found growing adventively. From these sites, five invasions of reproducing populations were identified and investigated further. These five sites, located in Bronx County, New York; Fairfield County, Connecticut; Philadelphia County, Pennsylvania; Queens County, New York; and Tolland County, Connecticut, were then investigated for similarities in their forest composition that may help to explain forest susceptibility to invasion by P. amurense. This work reports that P. amurense is a much more widespread invader than previous literature suggests, and that the invasion is not limited to any particular forest type. The five main invasion sites examined here do not share the same top species in importance value calculations, and correspondence analysis shows little similarity between four of the five sites. Tree density, which ranges from 55 to 643 trees per hectare, is also shown not to affect P. amurense's ability to invade a particular site. These data provide strong evidence of the need for close monitoring and potential removal of P. amurense before populations in the region reach a point at which removal will no longer be an option.
Keywords: Invasive species, New York, Philadelphia, Connecticut, New Jersey, importance values, Phellodendron amurense, Rutaceae
Introduction:
Since publication of the landmark work The Ecology of Invasions by Animals and Plants by Charles Elton (1958), increased attention has been paid to the problems associated with non-native invasive species. More recently, the monetary cost of these invasions has begun to be examined, with some estimates as high as $137 billion per year (Pimentel et al. 2000). In spite of more than 50 years of research and potentially staggering costs, new species continue to be introduced into the United States, either intentionally or accidentally, on a regular basis. While the pace of research has increased in the past decade, with entire journals now dedicated to the study of invasions, relatively little is known about the vast majority of invasive organisms, including invasive vascular plants.
In the regions surrounding New York City from Boston to Philadelphia, numerous invasive plants have been well documented and researched, including Acer platanoides (Fang 2005), Ailanthus altissima (Knapp and Canham 2000), Lythrum salicaria (Blossey et al. 2001), and Berberis thunbergii (Ehrenfeld 1997). Numerous other species, including Viburnum lantana and Spiraea japonica, show invasive potential in the region (Martine et al. 2008) and will likely receive closer monitoring for invasion in the coming years. One species of particular interest here is Phellodendron amurense Rupr., commonly known as the Chinese or Amur cork tree. A dioecious member of the Rutaceae, P. amurense is proving to be a relatively widespread invasive plant throughout this region. In New York City, Glaeser and Kincaid (2005) reported that P. amurense ranks third in overall dominance based upon importance value within a woodland, showing the need for further examination of the species. In spite of its prevalence, there is still very little recognition in the literature of the invasive potential, spread, and ecological impact of this species.
Phellodendron amurense is currently grown in the northeastern United States for its ornamental value. As a mature specimen, P. amurense has compound leaves with bright yellow fall color and abundant black fruits which are distributed by local birds, particularly the American robin, Turdus migratorius (Glaeser and Kincaid 2005). Dirr (1990) describes P. amurense as an excellent tree for parks and other large areas. A native of Asia, Phellodendron amurense, which is very cold tolerant, has apparently never established itself as a major horticultural species, as it is currently available only through specialty nurseries and mail order. This is possibly due to its large adult size of 10 to 15 meters in height and potentially greater spread, as described by Dirr (1990). The lack of horticultural availability makes the appearance of reproducing populations of this plant even more important to monitor, since they can easily be overlooked until long after they are unable to be removed. The need for early identification, monitoring, and removal of potentially invasive plants is often difficult to convey to professionals in the nursery trade. In a recent study of nursery professionals in Connecticut, 49% of those surveyed still worked with Acer platanoides and 66% with Berberis thunbergii (Gagliardi and Brand 2007), both of which are well documented and notoriously invasive plant species with major environmental impacts (Wycoff and Webb 1996, Ehrenfeld 1997). Early recognition of invasive tendencies in a particular species can prevent later problems trying to control it. When invasive species are already well established in both the natural landscape and the nursery trade, control is nearly impossible. This paper presents evidence that P. amurense is established over a large range in the New York City region, is capable of establishing in a wide variety of habitats, and warrants aggressive removal policies before it becomes a major problem in our region.
Methods:
In the summer of 2008 and 2009, field surveys were performed throughout Connecticut, and around New York City, Long Island, New Jersey, and Philadelphia to locate spontaneous, non-horticultural populations of P. amurense. These sites were selected based upon author observations, herbarium records, literature reviews, and the results of a request for sighting information which was distributed in early 2008 to botanical organizations and universities. Over 20 sites were documented, 14 of which were found to contain reproducing populations of P. amurense with both male and female trees, and these were analyzed further to determine the extent of the invasion. Five of these fourteen reproducing sites had at least five mature trees and were then analyzed to determine the types of surrounding vegetation in which P. amurense had established itself. These five sites, the Bartlett Arboretum, Fairfield County, Connecticut (41.07N 73.33W); Forest Park, Queens County, New York (40.42N 73.50W); Goodwin State Forest, Tolland County, Connecticut (41.46N 72.05W); Morris Arboretum, Philadelphia County, Pennsylvania (40.05N 75.13W); and the New York Botanical Garden, Bronx County, New York (40.51N 73.52W), were all surveyed further in 2009.
Each site was analyzed to calculate importance values (Curtis and McIntosh, 1951) for the surrounding vegetation. To perform this analysis, a variation of the Point-Center-Quarter (PCQ) method was used as follows. Transects were placed through the area invaded by P. amurense creating the longest route through the invaded area at each location. An additional transect was placed at both the Bartlett Arboretum and the New York Botanical Garden to sufficiently sample the larger invasion sites. Points were then chosen by using the closest P. amurense tree within 10 meters of the transect and over 5 centimeters dbh (diameter at breast height) as point one. The dbh for each tree closest to point one in each quarter was then recorded with its species identification and radius to the center point. If the closest tree in the quarter was another P. amurense, the second closest tree was sampled, and so forth. If no tree other than P. amurense was found within 20 meters of the center point, the species determination was listed as open, to represent an opening in the forest. The radius for that value was recorded at 20 meters. Point two along each transect was chosen by selecting the next P. amurense greater than five centimeters dbh within ten meters of the transect which did not have any tree in a quarter overlapping a tree in the previous PCQ data set. This process was repeated until no more data sets could be collected without overlap. A total of 27 different tree species was recorded at all five sites combined.
Importance values were calculated as described by Curtiss and McIntosh (1951). P. amurense individuals acting as the centroid points were not included in calculations of importance values. A bootstrap method in JMP 7.0 created by Kincaid (2008) was used to create an upper boundary importance value and a lower boundary importance value representing 95% confidence intervals. To determine the density of woody plants within the surrounding area, a bootstrap method by Kincaid (2008) was performed in JMP 7.0. Correspondence analysis was done using JMP 8.0 by analyzing the species by site occurrence matrix.
Results:
Site visits revealed that there are at least 14 sites in the region where P. amurense has successfully invaded. An additional 8 sites were found by searching the databases of several herbaria and reviewing the literature. These results are summarized in Table 1. Sites of invasion may have from several individuals to over 350 individuals. All sites had more than one individual, although in several cases only one tree greater than five centimeters dbh was found, and it wasn't possible to confirm existence of a female plant to ensure continued reproduction.
Importance values (IV) calculated for the five main sites of invasion reveal vast differences between the sites. With open space being calculated as a taxon value, as is done here, there was no perfect similarity among the highest-ranked taxa of the five sites. The New York Botanical Garden and Forest Park, which are also the two closest sites geographically, share four of the top five taxa in importance. However, Quercus rubra L. ranks first at Forest Park with an IV = 130.6, while at the New York Botanical Garden Q. rubra ranks third (IV = 38.6), and Prunus serotina Ehrh. ranks first (IV = 75.3). The additional three sites are all led in importance value by different species: the Bartlett Arboretum by Acer rubrum (IV = 93.1), Morris Arboretum by Acer negundo L. (IV = 141.4), and Goodwin State Forest by open space (IV = 143.8). These results are summarized in Table 2.
Calculations of absolute density of forest trees also show a vast difference in the surrounding vegetation of the invaded sites. The radius measurements taken between the centroid P. amurense at each point on the transect and the nearest non-P. amurense revealed that the radius ranged from 3.9 meters at the Bartlett Arboretum to 7.8 meters at Goodwin State Forest. A bootstrap resampling of these data revealed that the average density of non-Phellodendron trees at each site ranged from a low of 55 trees per hectare at Goodwin State Forest to 643 trees per hectare at the Bartlett Arboretum.. Bootstrapping to obtain minimum 95% confidence interval and maximum 95% confidence interval density values for each site revealed variation in these values; however, only the New York Botanical Garden and Forest Park produced values which could potentially overlap. These results are reported in Table 3.
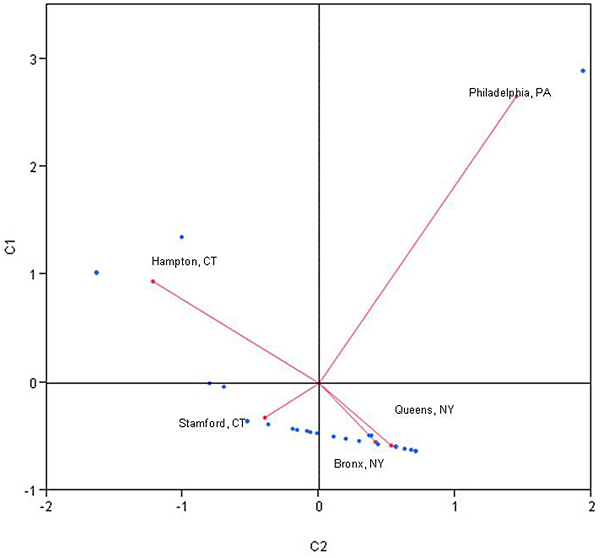
Correspondence analysis confirms the data from importance value calculations with more insight into site differences. The New York Botanical Garden and Forest Park are the most similar sites to each other, and along with the Bartlett Arboretum forest, are all most closely aligned with the centroid. Morris Arboretum and Goodwin State Forest show the most deviation from the centroid.
Discussion:
The extent to which P. amurense has invaded the region is much greater than previously reported. Gleason and Cronquist (1991) report P. amurense as occasionally escaped, and Lamont and Young (2002) report P. amurense as occurring at only two sites within the approximate range of this work. Factors influencing the invasiveness of a species can be numerous and difficult to disentangle. At the Bartlett Arboretum site, deer herbivory is common on a variety of woody and herbaceous species; however, browsing by deer has never been noticeable upon P. amurense at this site in more than three years of various studies on the species. This potential factor could explain part of its success. However, at both the New York Botanical Garden and Forest Park sites, deer are rare to nonexistent. More intriguing is the trees' potential impact on surrounding species through either competition or other factors such as potential allelopathy. These and other potential aspects of P. amurense's success will all need to be further examined.
This work documents numerous sites, including several which comprise hundreds of individual plants. It also shows that P. amurense is invading a range of forests throughout the region studied and has invaded both open areas and established woodlands. Yoshida and Kamitani (1999) describe P. amurense as a shade-intolerant species in the forests of central Japan; however, the evidence presented here contradicts their observations of the plant in its native range. These discrepancies can be due to a number of factors. The surrounding species composition in the plant's native range may affect P. amurense in ways that the regional flora in its invaded area cannot; for example, times of leaf emergence and leaf drop might be longer or shorter for species surrounding P. amurense in either site. Additional possibilities include the effects of apparent variability within the species and the potential for hybrid vigor between varieties of this species. Williams et al. (2005) showed that in Schinus terebinthifolius Raddi, an exotic invasive species in Florida, hybridization between two distinct introductions of the plant resulted in a hybrid variety that is the genotype of the plant spreading across the state of Florida. As in the case of S. terebinthifolius, P. amurense has well-documented varieties within the species. Ma et al. (2006) revised the genus Phellodendron to contain only two species, P. amurense and P. chinense. Prior to this revision, the genus contained over ten species and numerous other varieties, based upon collections within their native range. Several studies (Greller 1977, Bertin 2005) within the geographic range of this paper have reported former names for P. amurense, possibly due to the apparent variation within the species that led to the original splitting of the genus. In an earlier study of Phellodendron within the New York Botanical Garden (De la Cruz and Nee 2003), specimens collected among horticultural and escaped individuals showed variation in both leaf-base shape and hairiness of leaf undersides. De la Cruz and Nee also document three separate plantings at the New York Botanical Garden originally determined as three separate species, P. amurense, P. japonicum, and P. sakhaliense. All three of these are now P. amurense according to Ma, increasing the chances that hybridization between variations in a single species formerly regarded as several distinct species may indeed play a role in its successful invasion. More detailed analysis using molecular techniques will determine the influence of this hybridization, if any, in the future.
The calculation of importance values for the five sites reveals a difference in the surrounding forest types of each invaded site. The results reported here show that the five sites have different dominant species, with Goodwin State Forest being characterized by open land. Goodwin State Forest is in the process of removing the P. amurense from the main invasion site, allowing access to tree rings of many trees. All the P. amurense specimens with rings available for counting were aged 27‒29 years. These dates coincide with a documented late spring storm which destroyed a large stand of pines (species unknown) 29 years before the survey of the site (Goodwin State Forest interpretive signage). This indicates that P. amurense almost immediately colonized this open space after the disturbance. The invasion history at other locations is less clear. Morris Arboretum, which is dominated by large individuals of Lonicera tatarica and Acer negundo, also has a significant amount of open space, and with the lower-growing species heights of both L. tatarica and A. negundo, could potentially be seen as a relatively open space as well.
The remaining three sites yield more intriguing results. At the Bartlett Arboretum forest, Acer rubrum dominates the forest, which has a mean density of 643 non-P. amurense trees per hectare. This density, along with a mean dbh of 21.8 centimeters, indicates an established, mature forest which has been invaded. The forests of the New York Botanical Garden, with a mean density of 367 trees per hectare and a mean dbh of 20.4 centimeters, and Forest Park, with a mean density of 307 trees per hectare and a mean dbh of 28.9 centimeters, also provide evidence that P. amurense is invading not only open habitats but well-established forests of different densities as well. Unlike the Bartlett Arboretum site, the surrounding forest of the New York Botanical Garden is dominated by Prunus serotina, and Forest Park is dominated by Quercus rubra. The dominance by three different species at these three sites provides evidence that P. amurense is capable of invading a variety of ecological niches in addition to different levels of forest establishment.
The correspondence analysis performed for the five sites provides further evidence of the differences between sites. Only the New York Botanical Garden and Forest Park fall within the same quadrant of importance value data, meaning that four of the top five species are the same for these sites, although in different orders of importance. The other three sites fall into separate quadrants, with the Morris Arboretum and Goodwin State Forest showing the greatest deviation from the centroid.
Conclusions
This study shows that the invasion of P. amurense in the region of New York City is much more widespread than earlier works have reported and that P. amurense is capable of establishing itself within a large range of forests of the eastern United States. While additional data are needed to determine the exact current distribution, it is likely that with better field recognition of the species, it may be found in many additional parcels of forest throughout the region. As with many invasives, early detection and eradication are important, and P. amurense presents itself as a candidate for invasive removal programs and designation by both government and nongovernmental organizations as an invasive or noxious weed.