
Butterfly Activity in a Residential Garden
by Christopher Young
School of Applied Sciences, University of Wolverhampton, Wulfruna Street, Wolverhampton, WV1 1SB, United Kingdom
Abstract
Butterflies are a highly visible, well-loved, and well-studied part of Britain's native fauna, yet there is still very little known about how butterflies use one of the country's most commonly available habitats, the residential garden. Studies in a Wolverhampton (UK) garden demonstrate that the majority of individuals use these spaces as movement routes through the urban matrix. Of 516 observed individual visits by butterflies over three recording seasons (2000–2002), only 13.8% involved a stop for some purpose. The duration of these visits was characteristically short, with a mean visit time of nine seconds. Individuals tended to fly through the study garden using distinct entry and exit points largely dictated by variations in structure within the study garden and in the immediately surrounding gardens. Individual garden use by butterflies would therefore seem to be defined as much by structural imperatives as by availability of nectar- or food-plant species. When considered as systems of interconnected green spaces on the level of the housing block (defined as a continuous area of residential land use bounded by infrastructure or contrasting land uses) and of the urban area as a whole, residential gardens represent an extraordinarily valuable and dynamic component of the urban habitat.
Keywords: residential garden, housing block, butterfly, flight paths, vegetation structure, corridor, urban green space
Click image to enlarge

Red Admiral feeding on hemp agrimony (Eupatorium cannabinum). Photo © C. Young.
Background: Butterflies and Gardens
Butterflies are one of the best-known and most charismatic groups of fauna (Asher et al. 2001; Dover and Sparks 2000; Vickery 1995). Encounters with these easily identifiable creatures, along with garden birds, are among the first real contacts many people have with wildlife, and the careers of many keen professional and amateur naturalists have been inspired by such sightings. There is, as a result, a thriving network of ecologists studying these creatures, including many who contribute to a wide range of research studies into the spatial distributions and population dynamics of both rare and common butterfly species. This research encompasses a variety of survey activities, from national schemes—for example, the Millennium Atlas of Butterflies in Britain and Ireland (Asher et al. 2001) and long-term monitoring programmes (UK Butterfly Monitoring Scheme 2006)—to studies of characteristic species assemblages of particular habitat types (Croxton et al. 2005; Van Swaay 2002; Dover and Sparks 2000) and metapopulation studies of spatially restricted or rare species (Heikkinen et al. in press).
Urban areas have come under closer investigation in recent times due to the recognition that conservation and management of urban habitats and species pose particular challenges (Angold et al. 2006; Young and Jarvis 2003; McDonnell et al. 1997). Because of their intrinsic appeal, generally known life-history requirements, and sensitivity to small changes in site conditions, butterflies have often been a significant part of these studies (e.g., Wood and Pullin 2002). Indeed, some larger-scale studies have focused wholly or partially on butterflies as indicators of urbanization and/or the effects of urbanization (Bock et al. 2007; Vanreusel and Van Dyck 2007; Blair and Launer 1997; Ruszczyk and De Araujo 1992). Despite this large research community, there appears to be relatively little research—with the exception of the recent BUGS (Biodiversity in Urban Gardens in Sheffield) Urban Regeneration (URGENT) project (Thompson et al. 2003 et seq.)—into the most common, everyday, urban garden habitat, even though it is recognized in many local conservation strategies (for example, Wildlife Trust 2000).
Click image to enlarge

Gatekeeper feeding on late season meadowsweet (Filipendula ulmaria). Photo © C. Young.
Gardens are a significant proportion of the fabric of urban areas in the UK, comprising 19–27% of land use (Smith et al. 2005), and constitute a significant area of extensive interconnected green space (Mathieu, Freeman and Aryal 2007). Yet we still know surprisingly little about their landscape or ecological roles (Smith et al. 2006; Chamberlain et al. 2004; Owen 1991). Although schemes such as the UK annual Butterfly Conservation Garden Butterfly Survey use data from amateur recorders to monitor gross nationwide trends in species, no survey has explicitly investigated "the mobility or duration of stay of butterflies in gardens. There appears to be a dearth of published work on this aspect." (Vickery 1995, 134).
When undertaken, movement studies on butterflies have tended to look at the regional movements of certain species (Binzenhofer et al. 2005; Schneider and Fry 2005; Pryke and Samways 2001) as well as landscape-scale interpopulation movements (Sutcliffe et al. 2003) and the resultant effect on genetic diversity (Wood and Pullin 2002). Although there have been some smaller-scale movement studies that have investigated the influence of minor landscape features, either experimentally (Haddad 2000) or in the wider rural landscape (Cant et al. 2005), very few have investigated the microscale—the features that determine whether an individual butterfly will move through the structural complexity at the scale encountered within an individual patch (Dover and Fry 2001; Sutcliffe and Thomas 1996; Loertscher, Erhardt and Zettel 1995; Dennis 1986). No studies have looked at this in relation to the garden habitat.
This study aims to rectify this omission by quantifying indicative residence times and overall garden use by butterflies in a residential British garden. The data set was collected over three recording seasons in a single garden, rather than via a spatially complete study, and as such it provides baseline data on garden use rather than on the distributions of species in gardens.
Study Site
Click image to enlarge
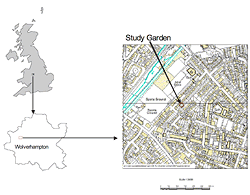
Figure 1: Location of the study garden within Wolverhampton, West Midlands, UK (© Crown Copyright/Database Right 2007. An Ordnance Survey/EDINA supplied service).
The study site is a residential garden in suburban northwest Wolverhampton in the West Midlands area of the UK (Figure 1). The 20-meter-long and 10-meter-wide garden is bounded on the north, south, and east sides by a 1.5-meter-high wooden fence and on the west side by a semi-detached house. The garden is adjacent to other gardens to the north, south, and east. Immediately bounding the garden is a variety of small garden buildings, trees, and shrubs (Figure 2a and Figure 2b). This includes sheds and garages that provide breaks of between 1 and 3 meters in the otherwise largely solid, shrubby boundary edge where it rises above the fence. There are also some larger and denser plantings with small gaps between adjacent canopies that act as barriers to the movement of wildlife. Within the garden, there is a similar mix of open areas and denser planting/solid structures (especially at the eastern end). The interplay of taller trees (> 5 meters), medium height shrubs (up to 5 meters), and the gaps between them resulting from garden management and garden buildings provides an intricate, though limited, network of potential routes for butterflies to use when moving in and out of the garden.
The study garden is one of approximately 100 such plots that form a continuous block of garden green space—which is itself one block among many thousands in the Wolverhampton urban area. In the context of this study, a "block" is taken to describe a continuous area of residential land use bounded by infrastructure or contrasting land uses and is used as a purely descriptive term. The garden itself is broadly wildlife-friendly, with a low input of artificial fertilizers, pesticides, and herbicides; apart from this, the garden is rather typical of urban UK gardens, being approximately rectangular in shape and consisting of a mixture of lawn, border flowers, herbs, shrubs, and trees.
Recording Butterfly Activity
To record butterfly activity, I used a hybrid of several established butterfly recording approaches. Recording time frames and environmental limitations were adopted from Pollard and Yates's (1993) standard Butterfly Monitoring Scheme (BMS) transect method. The practicalities of recording the movements of an individual visitor were adapted from the botanic garden method used by Wood and Samways (1991), and the method for recording and transcribing observations of butterfly behavior was adapted from Dover (1989).
Click image to enlarge
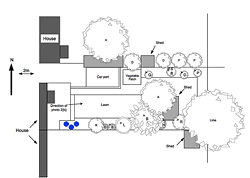
Figure 2a: Hatched areas show residential and garden buildings. Circular areas are significant garden shrubs or trees either rising above the surrounding fence or else with a strong attraction for butterflies, e.g., a series of buddleia bushes midway along southern boundary fence. Letters correspond to different species: Apple (A), Buddleia (B), Damson (D), Holly (H), Lilac (L) Plum (P), Quince (Q), Rowan (R) and Silver Birch (SB).
Click image to enlarge

Figure 2b: Looking east along the study garden (as marked on Figure 2a). Note the dense shrubs and trees at the far end and the high, dense barrier in the garden to the north and along parts of the southern margins.
To observe visitors, I undertook steady-paced, repetitive walks around the study garden. Flying butterflies were spotted prior to entering the garden and then were tracked over the course of their activity. The total range of each individual's activities was noted, and the timing of each activity, its flight track, and any stopping places were recorded as accurately as possible on a field recording sheet plan of the garden (Figure 2a). I did not actively hunt out butterfly visitors in the study garden's vegetation except when I tracked them there from flight. Each butterfly visit was recorded using a hand-held stopwatch, and times were rounded to the nearest second. Activities recorded were: flying, feeding, resting/perching, basking, and "other" (e.g., territorial displays).
The first individual observed was followed until it left the garden confines and airspace, even if it was still in sight (for example, moving into the neighboring garden), and any other individuals entering the garden during this time period were not recorded. If a butterfly flight was close to the garden edge and there was uncertainty as to whether it was just inside or just outside of the study garden, it was recorded as a visit. If a recorded individual left the garden, was kept easily in sight, and returned without alighting elsewhere—for example, took a simple flight path detour—its return was counted as part of the same visit to the garden (as in Dover 1989). Otherwise, return trips were counted as different visits because residence times, utilization of the garden, and activity during each visit were under examination, and not complete individual life histories. Butterflies were not recorded if their appearance was registered before and at the beginning of the recording period. However, if their ongoing activity coincided with the end of the scheduled recording period, butterfly visitors were tracked and recorded until their visit was completed.
The recording season extended from April 1 to September 29 (26 weeks), with one recording day attempted per week. Recording times were standardized to four hours each recording day with two hours during the morning (between 10:00 and 13:00) and two hours during the afternoon (13:00–16:00). Due to the sheltering effects of adjacent housing and the unique microclimates created by garden vegetation and artificial structures, and in an attempt to maximize the number of recording weeks, recording was undertaken in more marginal climatic conditions, i.e., when temperatures were slightly cooler than indicated in the BMS method, in the study garden than may have been attempted in "natural" habitats. As the recording period followed recommended BMS seasons, visits outside these recording months were not taken into consideration—although it is likely that urban gardens may have particular importance for butterfly movement and activity both early and late in the year, when nectar sources and fruit may be more available.
Laboratory and Statistical Analysis
The flight path of each butterfly was transcribed as closely as possible onto the recording sheet in the field, then transferred in the laboratory to the ArcView Geographical Information System to allow the investigation of trends and spatial use of the garden. The associated database was then exported to Microsoft Excel and the data analyzed for relationships using analysis of variance (ANOVA).
Click image to enlarge

Speckled wood perched on laurel (Prunus laurocerasus). Photo © C. Young.
General Results
Recording was undertaken in a total of 28 weeks out of a possible 78 over the 3 recording seasons of 2000–2002, 5 weeks of which produced no records. Twenty-five recording weeks were lost to bad weather and a further 25 to external commitments. Over the whole period, I recorded 13 species, out of a possible total of 59 native UK species. I also recorded a separate category of undetermined Small White species that flew through the garden quickly or at the farthest recordable distance and so were difficult to identify with certainty. Where these were recorded, I could not confidently identify these as Small Whites (Pieris rapae), Green-Veined Whites (Pieris napi), or female Orange Tips (Anthocharis cardamines), the "white" species recorded at other times in the study. In total, I recorded 516 individual butterfly visits in 112 hours of observation, with 278 visits in 2000, 128 visits in 2001, and 110 visits in 2002.
Two species dominated throughout: Large Whites (Pieris brassicae) and Small Whites (P. rapae) logged 169 and 138 visits, respectively. The least common visitors were Meadow Browns (Maniola jurtina) and Painted Ladies (Vanessa cardui), which registered only single visits. Commas (Polygonia c-album), Orange Tips (Anthocharis cardamines), Holly Blues (Celastrina argiolus), and European Peacocks (Inachis io) all also logged visit numbers in single figures, despite their traditional association with gardens.
Click image to enlarge
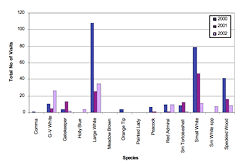
Figure 3: Number of visits to garden over all recording weeks by year, 2000–2002.
The three recording years show a variety of trends in both variability of recording time and butterfly numbers (Figure 3). The year 2000 had 15 recording weeks with 278 individual garden visits and a mean of 18.5 visits per recording session. Large and Small Whites dominated with 187 visits (>67%), while Meadow Browns, as noted above, showed only one record for all three seasons. The 2001 observation season saw 128 visits in 8 recording weeks (16.1 visits per session), with Large and Small Whites again dominant at 73 visits (>57%). The 2002 season had the fewest recording weeks at 5, but claimed the highest mean per session (22), with Large Whites comprising the majority (31.8%) of all observed butterfly species.
Garden Activity: Flight Times
Flight times of individual visits ranged between 2 and 128 seconds, with a mean visit time of 9 seconds. There was considerable variation among species, with Orange Tips having the shortest visit times, averaging 2.5 seconds, and Gatekeepers (Pyronia tithonus) having the longest mean visit times, at 16.7 seconds (Figure 4). Six species had mean flight times of between 5 and 10 seconds, and three further species had mean flight times between 10 and 11 seconds. The undetermined Small White species group had a mean flight time of 5.5 seconds, with a minimum of 3 seconds and maximum of 13 seconds.
Click image to enlarge
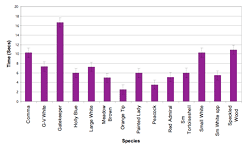
Figure 4: Mean flight times by species (±SE).
Despite the variety of timings recorded, the large variability within each species resulted in no significant differences in overall flight times between species (P=0.3154) or between years (P=0.2143). For species with more than 40 visits, there was some variability in flight times between years; both Small White and Speckled Wood (Pararge aegeria) showed no differences, while Green-Veined White (P=0.00005) and Large White (P=0.009) demonstrated significant year-to-year variability.
Garden Activity: Feeding, Basking, and Perching
Of the 516 individual visits, 71 involved stops for some purpose (13.8%). Fifty-two visits involved single stops, 14 showed 2 stops, and 5 visits had between 3 and 14 stops. Individual stops varied between 1 and 951 seconds, with feeding stops averaging 78.4 seconds and basking stops 99.2 seconds.
All individual visits to the garden were for a single purpose, mainly feeding or basking. The single exceptional visit involved five stops: three for nectaring, one for basking, and one for perching. Nine species stopped in the garden for some purpose, but only five species had two stops or more (Speckled Wood, Large White, Small White, Green-Veined White, and Red Admiral). The 27 recorded basking visits involved activity on such diverse substrates as a child's paddling pool, windowsill, and vegetation, while perching behavior was noted only on vegetation (no artificial perches) and during only eight individual visits.
Garden Activity: Flight Paths and Routes
Individual butterfly visitors exploited the garden landscape in a variety of ways, moving at a range of heights and responding both to the structural complexity of the garden surroundings and the internal heterogeneity of the garden (see Young 2005 for a fuller discussion of individual species responses). Despite this variety, there were significant uses of particular routes, with butterflies using common entry and exit points in response to a range of primarily structural modifiers of their behavior.
Click image to enlarge

Figure 5: All butterfly visit flight paths in the observation season of 2000. Distinct corridors of activity are noticeable both across and along the garden.
Gaps in the surrounding vegetation and hard structures both inside and outside the garden channelled flight paths. For example, individual visitors usually avoided any trees and shrubs that extended above the fence line, but used as entry or exit points the gaps created where neighboring sheds and garages extended above the fence line and as a result formed a break in the tree barrier (Figure 5). The trees in the garden to the north had a particularly noticeable influence on butterfly movement. They were substantially taller than those in the study garden and therefore created a channelling effect, as butterflies had to fly around a tree to exploit gaps between it and adjacent tree canopies and therefore deviated from otherwise straight flight paths both to and from the neighboring garden.
Missed Individuals
As this study was not designed to establish population sizes but rather to give an insight into activities during visits, it is certain that the results underestimate the number of visits made during the recording sessions and therefore overall. Ad hoc observations in 2000 and 2001 noted several individuals that were not recorded as they flew through while other visitors were being recorded. During 2002, an attempt was made to actively record numbers of these known missed individuals.
During week 7, when 39 visits were recorded, 9 further individuals were noted (>23% of total visits); during week 19, 39 were again recorded with 12 noted as missed (>30%); and finally, in week 24, 30 were recorded and 6 missed (20%). This indicates that the results underestimate the number of visits by 20 to 30%, especially considering that there were undoubtedly individuals missed but not noted.
As there were fewer individuals in total on the wing in spring and early autumn, it is likely that a greater proportion of the total number of butterflies using the garden was recorded in those seasons. In the summer months, several individuals were observed in the garden at the same time, with the inevitable result that a lower proportion of the total number was probably studied.
Discussion
As with any recording activity, the results of this study were heavily dependent upon when recording was possible. For example, the exceptionally poor early-season and late-season weather of 2001 resulted in low numbers of butterflies recorded in spring and autumn and restricted recording opportunities. None of the recording years had exceptional numbers of migrating vanessids except for late-season sightings of Red Admirals in 2000 and 2002 on warm, windless autumn days. Therefore, apart from the relatively low numbers of traditional garden species of butterflies, the species presences shown here are not especially noteworthy.
Click image to enlarge

Green-veined white feeding on marjoram (Origanum vulgare). Photo © C. Young.
Even allowing for both the underestimation of individual visits due to recording bias and the lack of studies with which to compare data, there were surprisingly small numbers of individual visits. Annual visit totals were usually boosted by a couple of busy recording weeks in midsummer. For example, in 2000, 87 individuals visited during one recording session (week 21), while one session had two visits (week 1), one had one visit (week 3), and three sessions had no visits (weeks 5, 7, and 22). There is also likely to have been a recording bias toward feeding and flying individuals because these are the most prominent (Dennis et al. 2006; Loertscher, Erhardt and Zettel 1995); therefore some resting, perching, and basking individuals could well have been overlooked.
The length of time that many butterflies spent in the garden was also unexpectedly low, with significant numbers, 47% of all visits, flying through in less than five seconds. It is evident that butterflies are using individual gardens as part of a wider meta-habitat, identifying available resources rapidly and then moving on if their requirements are not met at that particular time. In terms of determining nectar sources, for example, butterflies may be scrutinizing flower heads very rapidly (<1 second) (Dennis 1986; Goulson 2000), and this may have a commensurate impact on residence times.
The role of structural determinants of activity identified here reinforces the importance of 3D features in the landscape, especially the blocking and diversion effect of trees (Pryke and Samways 2001; Sutcliffe and Thomas 1996). Cant et al. (2005) suggest that butterflies adjust flight paths to avoid such features at distances of 100 to 200 meters, while Smith et al. (2005) emphasize how important boundary permeability is to the accessibility of gardens to wildlife. This suggests that butterflies are likely to avoid entering a residential block if the outside boundary is dense, or else the butterfly will have to make rapid internal responses to the structural variability that dictates its passage, which thus will influence its residence times and garden activity.
Conclusion
This study is based on data from one garden only, and as the layout, structure, and composition of any given garden are all very different, general conclusions may be difficult. However, it is clear that the variety of use patterns and timings noted here warrants further investigation across a wider range of gardens. Dover and Fry (2001) modified the behavior of species moving through agricultural landscapes by manipulating simple landscape structures; the data presented here suggest that it is likely that individual gardens can be similarly, and easily, managed to improve opportunities for butterflies to move through them and also to encourage them to stay longer. Manipulation could be as simple as ensuring gaps between adjacent gardens or planting relevant butterfly-friendly nectar and larval food plants. Such straightforward structural modification has been identified elsewhere as successful for bird species (Daniels and Kirkpatrick 2006) and is an inherent component of successful domestic gardening.
The data also suggest that individual visitors use the garden as a throughway rather than a stopping-off point. The short time spans recorded for individuals flying through indicate that, for butterflies, the garden functions as part of a route through the urban area within a wider garden landscape habitat, rather than as an isolated oasis in the local area (however "local" is defined). There were a number of distinct features that appeared to have an effect on the directionality of flight path, including garden orientation and the presence of shrubs, trees, and hard structures. Individual garden use would therefore seem to be defined by structural imperatives as much as by nectar or food plant species content, the advice characteristically given by wildlife gardeners to ensure that butterflies use a garden.
The consistently low numbers of individual visits recorded for much of the time were surprising. Single visitors dispersed throughout the day were either very visible or else showed small bursts of activity in response to rapid changes in weather conditions (e.g., periods of sunshine), and therefore contributed to the perception of a highly active butterfly garden, even if this was not the case most of the time. (Incidentally, a few very butterfly-active days in summer that coincide with times when the garden is well-used by people may well give a similar impression.)
However, when these relatively low visitation numbers are magnified to the level of gardens within the contiguous residential block (approximately 100 gardens in this instance), and then again to the level of the urban area as a whole, and finally across the UK, the importance of urban gardens to butterflies can be clearly demonstrated.
Acknowledgements
Thanks to Dr. Peter Jarvis and two anonymous referees for comments on earlier drafts.